
备案号:京ICP备2021034307号

概览

过去的十年里,类器官技术是干细胞研究中最重要的进展之一。类器官为三维的细胞培养物,重现了所代表器官的一些关键细胞类型和结构特征。这些「迷你器官」提供了高度相关的组织建模,从而使科学家可在体外研究复杂的学术问题。自从肠类器官1 和脑类器官2 的文献发表以来,类器官技术应用于多种物种和组织,可从成体干细胞(ASCs)或多能干细胞(PSCs)衍生构建。
虽然每种细胞培养系统都不同,类器官通常具有多种器官特异性的细胞类型,可重现所代表器官的功能。在类器官培养中,干细胞(PSCs 或 ASCs)在模拟体内信号环境的信号因子环境下进行培养。干细胞在该环境下增殖并分化为代表器官的细胞类型。总体来说,类器官具有与体内环境类似的结构,如顶侧和底侧表面的极化现象(上皮类器官),细胞分层现象(脑类器官)或复杂区域的重建(如小鼠肠类器官的隐窝绒毛结构)。在上皮类器官中,培养物中具有活跃增殖的成体干细胞,可以通过传代对培养物进行长期维持培养和扩增。
类器官已快速且广泛的应用于多项应用中,包括基础研究、转化医学和工业用途。类器官技术的根本在于发育生物学,所以类器官对其所模拟的组织提供宝贵的发育信息,尤其是 PSC 衍生的类器官。由于类器官重现了体内的信号通路和细胞间的相互作用,是研究细胞生物学中的宝贵模型,包括研究组织再生机制、干细胞 niche 的维持培养以及细菌、病毒和其他组织来源细胞的相互作用。
类器官具有其特有的实验优势,它去除了动物模型可能引入的混淆变量,但与均一化的细胞培养物相,则提供更高度的复杂性。通过结合高水平的生理相关性与体外操作的便宜性,在许多情况下,类器官具有辅助或替代体外使用原代细胞或永生化细胞系和动物实验的潜力。另外,类器官在培养中具有高度基因稳定性,维持了来源组织的基因型与表型。因此,类器官可进行疾病建模,用于研究疾病机制和进展,以及预测患者对药物治疗的个体化反应。
类器官方法的持续发展为干细胞研究带来了改革。在很多实验室,类器官已经用于互补现有的建模,用于开展曾经无法研究的方向。类器官培养技术的发展和标准化会使得类器官的可及性更广泛地实现。
肠类器官
肠类器官含关键性的肠细胞类型,包括肠上皮细胞、杯状细胞、肠内分泌细胞和肠干细胞,再现了肠上皮的生理结构。位于肠类器官内的活跃分裂的干细胞群保证这些类器官的长期稳定扩增和传代。
IntestiCult™ 类器官生长培养基(小鼠)和 IntestiCult™ 类器官生长培养基(人)分别支持小鼠 ASC 和人 ASC 衍生的肠类器官的构建和长期维持培养。STEMdiff™ 肠类器官试剂盒可用于从人多能干细胞直接分化为肠类器官3,通过 3 个阶段的分化,这些肠类器官可以在 STEMdiff™ 肠类器官生长培养基中进行维持培养和成熟。使用这些培养基可方便、灵活的构建实验所需的肠类器官,提高实验的稳定性,降低实验动物的使用。
表 1. 肠类器官培养系统的比较
肠类器官类型 | 小鼠(源于 ASC) | 人(源于 ASC) | 人(源于 PSC) |
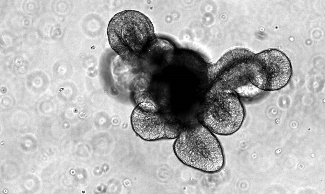
代表性图像 |
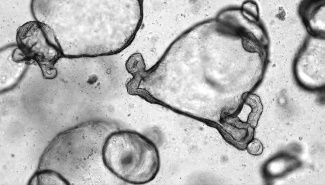
|
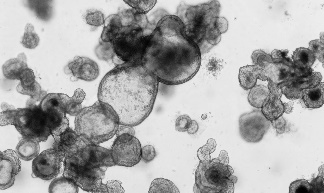
|
|
类器官细胞构成 | 类器官同时包括活跃的肠干细胞群和一群终末分化的细胞,重现了体内成熟的肠上皮系统。 | 经过 1-3 次传代的类器官主要包含处于肠祖/干样状态的细胞,类器官可后续在适宜的培养基中分化为成熟的肠道细胞类型。 | 类器官同时包括活跃的肠干细胞群,终末分化的肠道上皮细胞类型和相关的间充质细胞。类器官为胎儿表型。 |
起始样本 | • 小鼠小肠或结肠隐窝 • 小鼠肠类器官(产品号 #70931) | • 人小肠或结肠隐窝 • 已构的 ASC 来源的肠类器官 | • 人诱导多能干细胞(iPSC)或胚胎干细胞(ES) • 已构建的 PSC 来源的肠类器官 |
类器官维持培养 | 类器官可以被长期传代或冷冻保存。 | 类器官可以被长期传代或冷冻保存。 | 类器官可以被长期传代或冷冻保存。 |
遗传学考量因素 | • 已制定的健康和疾病模型 • 体内遗传操作的工具 | • 具有供体特异的遗传学背景 • 可体外进行基因编辑 | • 具有供体特异的遗传学背景 • 可体外进行基因编辑 |
适宜使用的培养基 | IntestiCult™ 类器官生长培养基(小鼠)(产品号#06005) | IntestiCult™ 类器官生长培养基(人)(产品号 #06010) | STEMdiff™ 肠类器官试剂盒(产品号 #05140) STEMdiff™ 肠类器官生长培养基(产品号 #05145) |
肝类器官
肝类器官培养技术的发展为肝脏研究领域提供了体外维持肝细胞持续培养的便捷方法4,5。肝祖细胞类器官的培养源于祖细胞的扩增,祖细胞被认为位于肝管内。肝祖细胞形成的球状类器官,主要由祖细胞构成,转移至分化培养基后,可进一步分化为肝细胞和胆管细胞6。
HepatiCult™ 类器官生长培养基(小鼠)是一种无血清的培养基,可以从小鼠肝脏组织快速生成肝祖细胞类器官。起始样本可来源于肝管、肝管小段、单个细胞或冷冻保存的类器官,并在 Corning® Matrigel® 液滴(domes)中培养,或在含有稀释的 Matrigel® 培养基中进行悬浮培养(suspension)。

图 1. 小鼠肝祖细胞类器官可来源于各种起始样本
无论是从(A)肝管小段(B)单个细胞或(C)冷冻保存的类器官,都可在 HepatiCult™ 类器官生长培养基(小鼠)中生成肝祖细胞类器官。所有的类器官在 Matrigel® dome 中生长,并在原代培养的第 7 天或解冻后的第 1 代(冷冻保存的类器官)成像。
胰腺类器官
胰腺类器官来源于从胰腺导管中分离出的胰腺干/祖细胞的扩增7。在添加有适当的生长因子和细胞外基质的培养环境中,胰腺祖细胞形成胰腺外分泌类器官。另外,从小鼠原代肿瘤和转移瘤中分离的组织可以构建出肿瘤源性的类器官,为胰腺癌和胰腺导管腺癌的研究提供模型。这些类器官保留了肿瘤的关键特征,包括基因表型和细胞表型,这为体外实验提供了一个方便的模型系统。
PancreaCult™ 类器官生长培养基(小鼠)适用于来源于胰腺导管、导管小段、单个细胞或冻存的类器官的胰腺外分泌类器官的生长。胰腺类器官的维持可以通过长期传代或冷冻保存,为实验提供稳定的细胞来源。

图 2. 小鼠胰腺类器官可来源于各种起始样本
PancreaCult™ 类器官生长培养基(小鼠)可从(A)胰管、(B)单个细胞和(C)冷冻保存的类器官中生成胰腺外分泌类器官。所有的类器官都在 Matrigel® dome 中生长。在原代培养的第 4 天或第 5 天(分别对胰管和单个细胞)或第一次解冻后的第 3 天(冷冻保存类器官)成像。
脑类器官
hPSC 衍生的脑类器官则是一种三维(3D)体外培养系统,能够重现人脑发育过程和发育中的人脑结构。这些「迷你脑」提供了一种模拟人体生理环境的体外模型,专用于研究人神经系统特有的神经发育和疾病过程。脑类器官在人脑发育和神经系统疾病(诸如自闭症、精神分裂症、或因寨卡病毒感染导致的脑缺陷)的研究中具有重要应用。
STEMdiff™脑类器官试剂盒为无血清培养系统,设计用于从人胚胎干细胞(ES)和诱导多能干细胞(iPS)生成脑类器官,基于 MA Lancaster 和 JA Knoblich8 发表的配方研发。培养 40 天后,这些脑类器官具有层次分明的祖细胞群,并产生成熟神经元,与在早期发育中人脑皮质层的观察结果相符。

图 3. STEMdiff™ 脑类器官试剂盒的示意图
将在 mTeSR1™ 中维持培养的人多能干细胞(胚胎干细胞或诱导多能干细胞)使用温和细胞解离试剂(GCDR)解离为单细胞悬液,以密度为 9000 细胞/孔接种至 U-Bottom 96 孔超低粘附培养板(Corning®)中,接种培养基为 EB 形成培养基 + 10 μM Rho-kinase 抑制剂(ROCKi)。每隔 2 天更换为不含 ROCKi 的 EB 形成培养基。5 天之后,将 EBs 转移到含有诱导培养基的 24 孔超低粘附培养板(Corning®)中。对 EB 培养 2 天后,使用液体 Matrigel®(减少生长因子,Corning®)包被 EBs。接下来将它们转移到经过无组织培养处理的 6 孔板(12-16 类器官/孔)中。嵌入的类器官在扩张培养基中维持培养 3 天。在第 10 天时,将类器官转换到成熟培养基中,然后放置在 RPM 为 57-95 的定轨摇床(Infors HT)上对其进行培养。在成熟培养基中,每 3-4 天进行换液。在第 40 天时,类器官进行 RT-qPCR 分析或冰冻切片后进行免疫染色。EBs 形成、诱导和类器官扩张的比例尺 = 300 μm。类器官成熟的比例尺 = 1 mm。
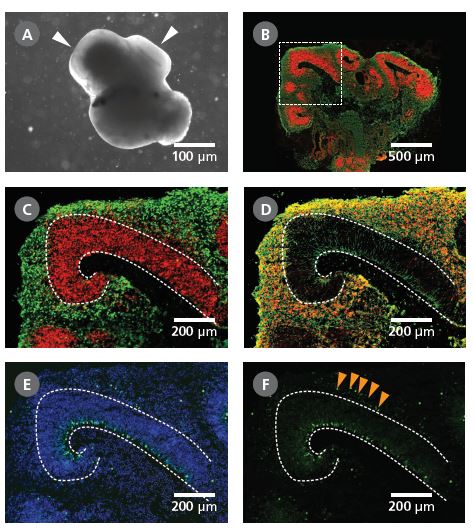
图 4. 对使用 STEMdiff™脑类器官试剂盒生成的脑类器官进行鉴定
(A)使用 STEMdiff™ 脑类器官试剂盒生成的整个脑类器官(第 40 天)于相差显微镜下的代表性图像。这一时期的脑类器官具有暗相衬结构,且该结构周围可能是更薄更透明的、显示分层的区域(箭头所示)。(B)在对脑类器官冰冻切片进行免疫组化分析时,通过顶端祖细胞标志物 PAX6(红色)和神经元标志物 III 类 β-微管蛋白(TUJ-1)(绿色),显示出类器官内部的皮质区域。(C-F)对(B)划线区域的放大图像。(C)PAX6+ 顶端祖细胞(红色,虚线之内)位于脑室带样区域。III 类 β-微管蛋白+神经元(绿色)位于该脑室带附近。(D)CTIP2(即发育中皮质板的标志物)与 III 类 β-微管蛋白+神经元共同位于皮质板样区域。各层结构重现了在人脑发育中观察到的早期脑皮质发育。(E)被 Ki-67(绿色)标记的增殖祖细胞沿脑室带分布,细胞核被 DAPI 复染(蓝色)。(F)在外脑室下区状区域中发现了额外的 Ki-67+细胞群(箭头所示)。比例尺:(A)1 mm、(B)500 μm、(C-F)200 μm。
产品信息
产品 | 规格 | 产品号 # |
IntestiCult™ 类器官生长培养基(小鼠) | 100 mL | 06005 |
小鼠肠类器官 | 200 个类器官 | 70931 |
IntestiCult™ 类器官生长培养基(人) | 100 mL | 06010 |
STEMdiff™ 肠类器官试剂盒 | 1 盒 | 05140 |
STEMdiff™ 肠类器官生长培养基 | 1 盒 | 05145 |
HepatiCult™ 类器官生长培养基(小鼠) | 100 mL | 06030 |
小鼠肝类器官 | 2 个培养孔 | 70932 |
PancreaCult™ 类器官生长培养基(小鼠) | 100 mL | 06040 |
小鼠胰腺类器官 | 200 个类器官 | 70933 |
STEMdiff™ 脑类器官试剂盒 | 1 盒 | 08570 |
STEMdiff™ 脑类器官成熟试剂盒 | 1 盒 | 08571 |
参考文献
1. Sato T et al. (2009) Single Lgr5 stem cells build crypt-villus structures in vitro without a mesenchymal niche. Nature 459(7244):262–5.
2. Lancaster MA et al. (2013) Cerebral organoids model human brain development and microcephaly. Nature 501(7467):373–9.
3. Spence JR et al. (2011) Directed differentiation of human pluripotent stem cells into intestinal tissue in vitro. Nature 470(7332):105–9.
4. Huch M et al. (2013) In vitro expansion of single Lgr5+ liver stem cells induced by Wnt-driven regeneration. Nature 494(7436):247–50.
5. Huch M et al. (2015). Long-Term Culture of Genome-Stable Bipotent Stem Cells from Adult Human Liver. Cell 160(1-2):299–312.
6. Broutier L et al. (2016) Culture and establishment of self-renewing human and mouse adult liver and pancreas 3D organoids and their genetic manipulation. Nat Protoc 11(9): 1724–43.
7. Huch M et al. (2013) Unlimited in vitro expansion of adult bipotent pancreas progenitors through the Lgr5/R-spondin axis. EMBO J 32(20): 2708–21
8. Lancaster MA & Knoblich JA. (2014) Generation of cerebral organoids from human pluripotent stem cells. Nat Protoc 9(10):2329–40.
该文章来自于网络转载,版权归原作者所有。如有涉及侵权请给予告知,我们将尽快删除相关内容。